
Internal anatomy
Skeleton
Skeleton
The internal skeleton of skates and rays (or endoskeleton) - as with sharks - lacks true bone, and is instead made entirely of cartilage. Cartilage is a strong and durable material that is lighter and more flexible than bone, enabling elasmobranchs (which lack a swim bladder) to stay afloat and turn in a tighter radius than other fish. However, parts of the elasmobranch skeleton - such as the skull, the vertebrae, and dermal spines - are often strengthened by the deposition of calcium and salts, a process called calcification. When the skeletal structures of elasmobranchs become sufficiently calcified, they can resemble bone, and have similar strength characteristics to bone, without the added weight. The process of calcification varies greatly from one species to the next, with some species - especially deep-sea and primitive forms - having little calcification while in others, such as pelagic sharks, calcification is extensive.
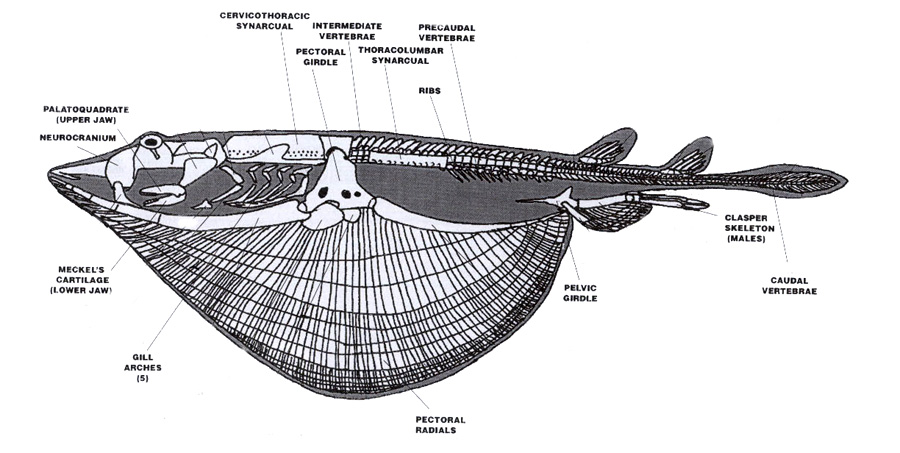
Diagram of an idealized batoid skeleton in lateral view showing the main skeletal structures. Photo modified from Sharks, Skates and Rays: The Biology of Elasmobranch Fishes. 1999. Edited by William C. Hamlett. John Hopkins University Press, Baltimore, Maryland.
The vertebral centra of elasmobranches are cylindrical and biconcave in shape and are generally divided into two primary types: precaudal and caudal vertebrae. The number of vertebrae a given individual will contain throughout its entire life is set during embryonic development, a period which also marks the beginning of vertebral calcification. Precaudal vertebrae generally begin forming before caudal vertebrae, and all vertebrae appear to grow throughout the lifespan of individual sharks, skates, and rays. This enables the use of vertebral centra for ageing. Age is determined in elasmobranchs by counting vertebral rings that are deposited annually, not unlike those of trees. This method is analogous to the use of otoliths (or earstones) to determine age in bony fishes. For more information on how skates are aged, so to the Skate Research section of this website.
In batoids, the spinal column is quite elaborate compared with that of sharks. Specifically, posterior to the neurocranium (or the elasmobranch skull) at the anterior end of the vertebral column, the precaudal vertebrae are fused into a tube called a cervicothoracic synarcual, which can include as few as 4 and as many as 45 vertebrae, depending on the species (Fig. 1). This rigid structure is much less flexible than the rest of the vertebral column, and functions to support the greatly expanded pectoral fins. Some species of myliobatoids even have a 2nd synarcual!

Ventral view of an embryonic thorny skate (Amblyraja radiata) viewed under reflected light. The cartilaginous pectoral fin radials extending to the distal edges of the right pectoral fin are clearly visible.
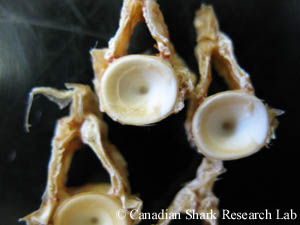
Thoracic vertebrae removed from a thorny skate (Amblyraja radiata)

A great hammerhead (Sphyrna mokarran) vertebral centra. The growth rings are clearly visble.
Liver
Liver

A mature female thorny skate (Amblyraja radiata) dissected to reveal the large, lobed liver (which has been removed from the body cavity). The liver is the first organ encountered when a ventral incision is made in the body cavity.
The liver of elasmobranchs is a large, soft and very oily organ which occupies most of the body cavity and can comprise as much as 25% of the body weight. It is the first organ encountered when an incision is made ventrally from the axils of the pelvic fins to the anterior end of the body cavity. It consists of two large, pointed lobes that are greenish-gray to dark reddish-brown in colour. The function of the liver in sharks, skates, and rays is two-fold. First, as in all animals, the liver concentrates the fatty reserves, and therefore provides for energy storage. Secondly, the liver of elasmobranchs acts as a hydrostatic organ by storing lighter-than-water (or low density) oils. These oils counteract the sinking tendency of the elasmobranch by decreasing the density and increasing the buoyancy of the animal on a whole, such that the animal becomes neutrally buoyant. Without such a large liver, sharks, skates and rays would have difficulty staying off the bottom, as they lack the swim bladder characteristic of bony fishes.
Digestive Tract
Digestive Tract
The second most noticeable structure in the body cavity of sharks, skates, and rays is the digestive tract which consists of two contiguous organs: the esophagus and the stomach. The anterior end of the stomach (also known as the cardiac stomach) is J-shaped and saclike, and tapers into the posterior part of the stomach known as the pyloric stomach, which bends anteriorly. The pyloric stomach terminates at a constriction called the pylorus, which leads to the short duodenum and then to the larger spiral valve intestine, which is highly coiled and twisted internally. The function of the spiral valve intestine is to increase the surface area for digestion and absorption of food, while also conserving space in the body cavity for the large liver and for the development of embryos (in the case of rays) or egg capsules (in the case of skates). The spiral valve, in turn, leads to the rectum and the anus, which opens into the cloaca, a cavity where the digestive, urinary, and genital glands open to the outside.
Pancreas
Pancreas
The pancreas is a gland that aids in digestion by secreting digestive enzymes into the duodenum. It consists of two connected lobes: a ventral lobe, which contains a duct from which pancreatic secretions enter the duodenum, and a dorsal lobe. Both are usually pinkish in colour.
Spleen
Spleen
The spleen is a dark brownish organ, triangular or slightly elongate in shape, which lies against the stomach. However, it does not play a role in the digestive process. Instead, it is part of the lymphatic system, a system which is a major component of the immune system.
Rectal Gland
Rectal Gland
The rectal gland is a small, finger-like organ that concentrates large quantities of excess salt (or sodium chloride) from the bloodstream for eventual excretion via the anus. Specifically, it secretes a colourless solution with about twice the concentration of sodium chloride found in the blood plasma into the rectum via a small duct. This organ is very important to elasmobranchs, whose livers produce large amounts of urea, thereby making these marine fishes slightly hyperosmotic to seawater (i.e. their internal solutes are greater than the surrounding seawater, so they are constantly gaining water and salt).
Kidneys
Kidneys
The kidneys are part of the urogenital tract, and are involved in the manufacture and transport of urine as well as in the regulation of plasma urea concentrations. In skates and rays, they are either semi-lunar shaped or ribbon-like, dorsoventrally flattened, dark red organs that are highly lobed and lie dorsally on either side of the spinal column outside of the body cavity. A tough membrane, called the peritoneum, separates the kidneys from the rest of the body cavity. The kidneys are drained into the cloaca by the ureters.
Male Reproductive Tract
Male Reproductive Tract

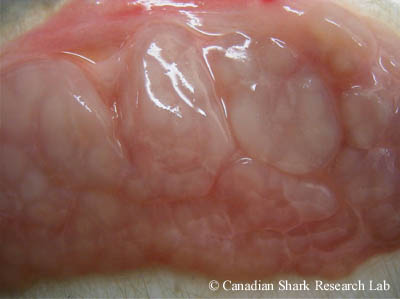
An immature male smooth skate (Malacoraja senta) cut open to reveal the whitish-yellow testes with indistinct lobes. Note the small, undeveloped claspers.
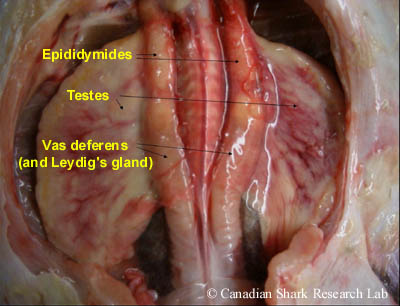
The organs of the male urogential tract include the male gonads (or testes), genital ducts, the Leydig gland, and the alkaline gland. Removal of the digestive tract exposes the paired, flattened testes, which are whitish or yellowish in colour and are at the anterior end of the body cavity dorsal to the liver. In many species, the testes are enveloped in an organ called the epigonal organ, which is thought to play a role in blood-cell formation. Each testis is an aggregation of irregularly-shaped lobes; in immature sharks the testes are an inconspicuous mass of whitish tissue with indistinguishable lobes, whereas in mature sharks the lobes are distinct and bulging.
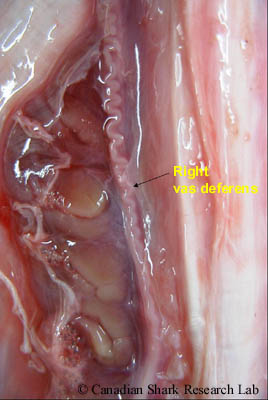
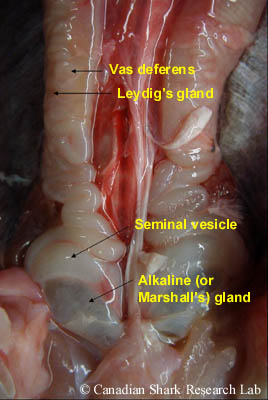
Sperm cells (or spermatocysts) are produced and develop in the lobes of each testis (a process called spermatogenesis). Mature sperm (spermatozoa) are then conveyed through minute ductules (called ductus efferens) into a highly coiled epidydimis, which can be seen along the vertebral column on either side of the dorsal aorta. Continuing posteriorly, the sperm pass from the epidydimis into the ductus deferens (or vas deferens). This is where sperm are stored prior to copulation, and also where spermatophores are formed. Spermatophores are ovoid or spherical bundles consisting of a large number of aggregated sperm that function to protect the sperm and to prevent leakage and loss of sperm into the water during mating. Both the epididymis and vas deferens receive a viscous secretion produced by the adjacent Leydig gland which is thought to stimulate the movement of spermatozoa.
In immature males the vas deferens is a thin, straight tube, whereas in sexually mature males the anterior part of the vas deferens is highly convoluted and sinuous, covering most of the kidneys. Immature and mature males are also distinguished by the posterior part of the vas deferens, which in mature males is greatly enlarged to form the seminal vesicles. On the ventral side of each seminal vesicle is an outpocketing (or lateral dilation) termed the sperm sac; the two sperm sacs unite posteriorly to form the urogenital sinus that ends as the urogenital papilla emptying into the cloaca. The name "sperm sac" is actually a misnomer, as these structures do not store sperm. In various species of skate, these structures are actually a specialized gland (termed the alkaline or Marshall's gland) containing a fluid of high alkalinity that neutralizes the acidic urine and may also be involved in sperm protection and/or the formation of copulatory plugs. The cloaca is located between the pelvic fins and is common to both sexes. It is the chamber where the digestive, urinary, and genital glands open to the outside.
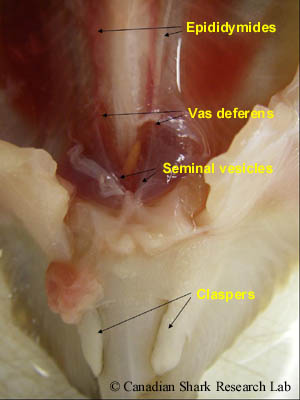
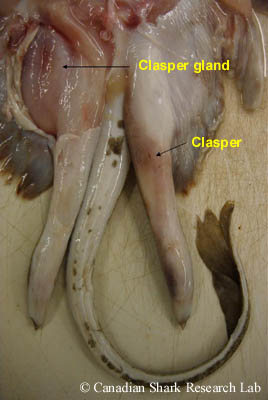
In male elasmobranchs, the inner margins of the pelvic fins are modified to form paired, tube-like copulatory appendages known as myxopterygia or - more commonly - as claspers. During mating the claspers are rotated forward, one is inserted into the female, and sperm (or spermatophores) travel down the dorsal, longitudinal groove of the clasper into the cloaca of the female. In sharks, the sperm are forcibly ejected from the cloaca down the clasper groove by a muscular, contractile organ called a siphon sac located beneath the skin on the ventral side. Specifically, prior to mating, seawater is pumped into the sacs by flexion of the claspers and then ejected when the siphon sacs contract, washing sperm into the female.
In batoids, siphon sacs are not present; however, there are paired, ventrally situated clasper glands ensheathed with striated muscle that project into a clasper sac near the pelvic fins. When the striated muscle is contracted, secretions from the clasper gland are forced into the clasper sac, which then forces secretions down the clasper groove where it is mixed with sperm from the cloaca. Gland secretions act as a lubricant and a transport medium for sperm, and also seal the clasper groove to prevent dilution of the semen in seawater and sperm escape. Mating in batoids is a prolonged activity typically lasting approximately 1 to 4 hours; however, it is not frequently observed in any species of elasmobranch.
If you want more information on how maturity and reproductive cycles are examined in skates, go to the Skate Research section of this website.
Female Reproductive Tract
Female Reproductive Tract
In females, removal of the digestive tract exposes the urogenital tract, which consists of the gonads (or ovaries), the ostium, the oviduct, the nidamental (or shell) gland, the uterus, and the urogential sinus. The ovaries - which are whitish yellow in colour - are located at the anterior end of the body cavity, dorsal to the liver.
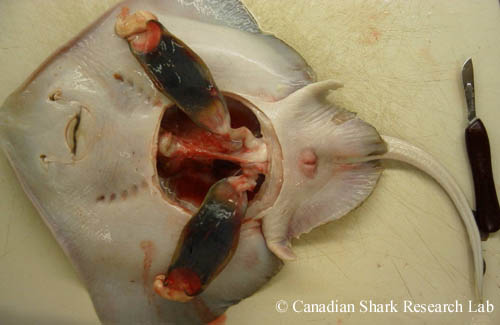
A mature female thorny skate (Amblyraja radiata) opened for examination of the reproductive tract. Two fully developed egg capsules are visible in the uteri below the shell glands.
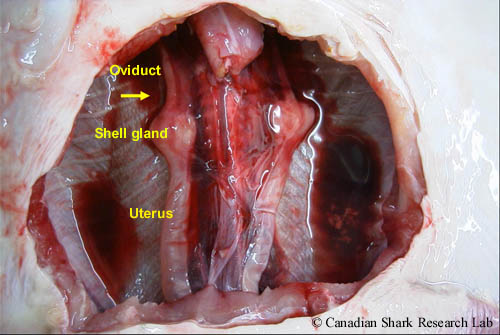
The reproductive tract of a maturing female thorny skate (Amblyraja radiata). The yellow arrow marks the location of the right ovary, which was removed to show the more dorsal reproductive structures.

The reproductive tract of a mature female thorny skate (Amblyraja radiata). Note the large, vitellogenic ova in her ovaries, the enlarged shell glands, the flaccid uteri, and the swollen urogenital sinus.
In immature females, the ovaries are thin and smooth in appearance. As the female matures, many small white oocytes start to become visible, embedded within the connective tissue of the ovaries, giving them a granulated appearance. In sexually mature females the ovaries are large, often bearing many yolked (or vitellogenic) eggs at the surface, in addition to the many small developing follicles of various sizes embedded within the connective tissue. In some species of elasmobranch, only one ovary is functional and contributes to egg production, as is commonly the case in rays where the left ovary is the principal generator of mature ova and the right is rudimentary. In skates, both ovaries tend to be functional.
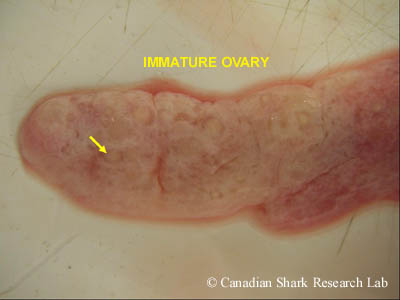
The ovary from an immature winter skate (Leucoraja ocellata). Note the small, indistinct whitish ova embedded in the connective tissue.

The ovary from an early- maturing winter skate (Leucoraja ocellata). Note the ova beginning to differentiate in size and colour.
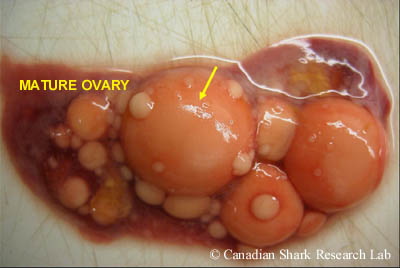
The ovary from a mature winter skate (Leucoraja ocellata). Note the very large, yellowish-orange vitellogenic ova ready to be ovulated, and the smaller ova of varying sizes and vitellogenic states.
Rays
In batoids with well-defined reproductive cycles, such as many species of rays, the beginning of the reproductive season is marked by an increase in the number and size of oocytes at the anterior surface of the ovaries (a process termed oogenesis). Oocytes also gradually begin to accumulate yolk (a process termed vitellogensis) - changing from whitish to yellowish-orange in colour - until they are ready to be ovulated. This process is slow, requiring several months to a year in most species. During ovulation, the largest mature ova are released one by one from the ovaries to the ostia (singular: ostium) and then into the oviducts, which are long tubes that run the length of the body cavity and whose sole function is egg transport. The release of mature eggs from each ovary stimulates other mature eggs to migrate to the periphery of the ovary surface, such that a continual supply of eggs is available for ovulation. After passing through the anterior oviduct, each ovum enters the nidamental (or shell) gland, which is an enlargement of the anterior end of the oviduct. The shell gland is an extremely specialized organ where sperm are stored (in some species of batoid); where fertilization occurs; and where each fertilized ovum is subsequently encapsulated in a thin, membranous egg case or "envelope" prior to parturition. The size of the shell gland correlates with the reproductive cycle, reaching a maximum just prior to the beginning of ovulation. Posterior to each shell gland is the uterus, an enlargement of the posterior oviduct. The two uteri unite to form the urogenital sinus, a chamber which opens into the cloaca and enlarges concomitant with sexual maturation.
In rays, mating, gestation and parturition tend to occur in synchrony. Follicle development in the ovaries can occur at the same time as pregnancy (Fig. 7) - which is typical of elasmobranchs with year-long gestations - or between pregnancies, which is the case in most rays with comparatively short pregnancies of several months within the yearly cycle (Fig. 8). The ovaries of reproductively inactive females (i.e. at the end of the reproductive cycle), contain only small follicles with little or no yolk. Gestation is followed almost immediately by mating and the ensuing pregnancy. Exceptions include Torpedo marmorata which has a year-long gestation and two non-pregnant years between pregnancies. Parturition, in turn, may be triggered by specific ecological factors, such as changes in temperature or relative abundance of prey for the neonates.

The right shell gland and anterior uterus of a mature Northwest Atlantic skate. In skates, the shell glands dominate the reproductive tract, whereas in rays, the size of the shell glands fluctuates noticeably with the timing of the reproductive cycle.
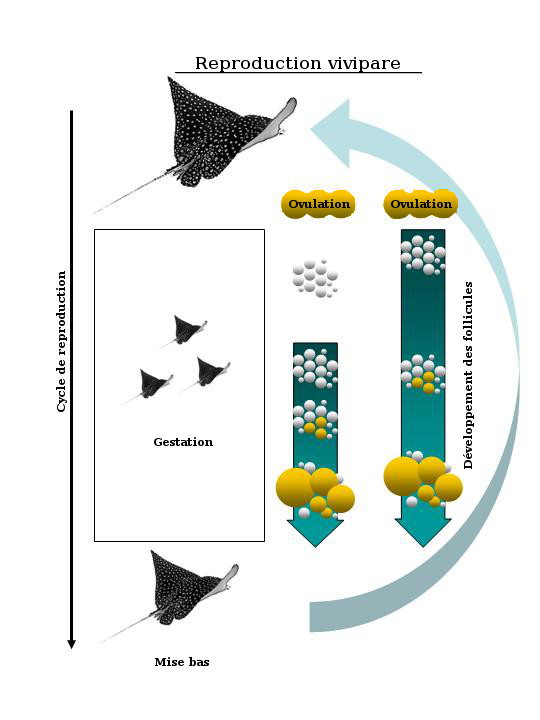
A schematic showing two viviparous reproductive cycles typical of rays with year- long gestation periods. In the first scenario, follicle development is occurring during the latter half of pregnancy (or gestation). In the second scenario, follicle development is occurring throughout pregnancy.
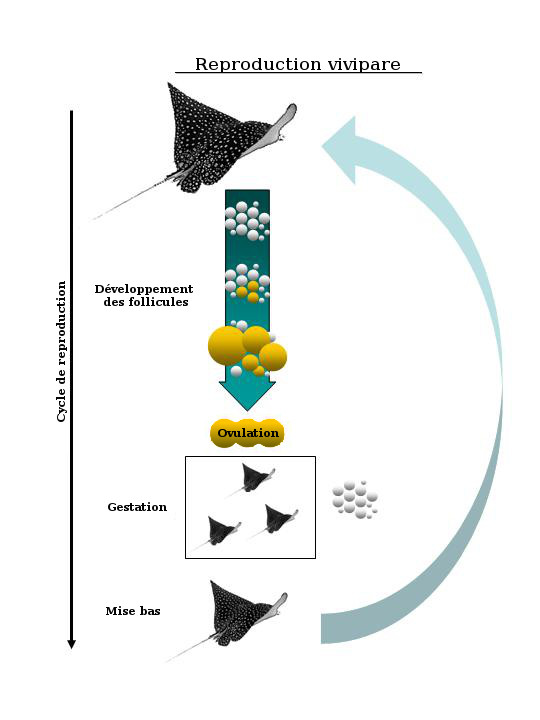
A schematic showing the viviparous reproductive cycle typical of rays with shorter gestation periods lasting several months within a yearly cycle. Follicle development occurs between pregnancies.
Skates
The internal reproductive anatomy of skates is very similar to that of rays. Unlike in rays, however, the seasonal reproductive cycles of oviparous species of elasmobranchs (including all species of skates) are generally not well-circumscribed with a clear beginning and end. Instead, the general trend is one of year-round egg production with seasonal periods or peaks when a higher proportion of the mature females are laying eggs. In order for year-round egg production to occur in these species, a graded series of paired follicles is present at all times within the ovaries, with each pair requiring at least several weeks to attain pre-ovulatory size. Vitellogenesis occurs in synchrony with oviposition. In other words, mature females exhibiting this "conveyer belt" type of reproduction have larger yolked ova and many small white ova of all sizes in their ovaries during every month of the year, with smaller eggs maturing constantly to replace those that are encapsulated and oviposited (Fig. 2).
Also distinct is the production of egg capsules by skates relative to that occurring in rays. In skates, as with rays, encapsulation of the ova occurs in the shell glands. However, in all oviparous species, the shell gland dominates the reproductive tract, with its size increasing coincident with periods of peak ova production, then fluctuating only slightly during the reproductive cycle. In addition, because of the complexity of capsule formation in oviparous species, formation begins prior to ovulation and is one third to one half complete when the fertilized egg enters the shell gland. The fully completed egg capsules produced by skates are structurally complex and remarkably durable, and are released into the external environment during a process termed oviposition. Egg capsule formation is, nevertheless, a quick process requiring only 12 -24 hours, with capsule hardening - or sclerotization - occurring in the uterus. Fully-formed egg capsules are then housed in the urogenital sinus for several hours prior to oviposition, which in turn, can take several days.
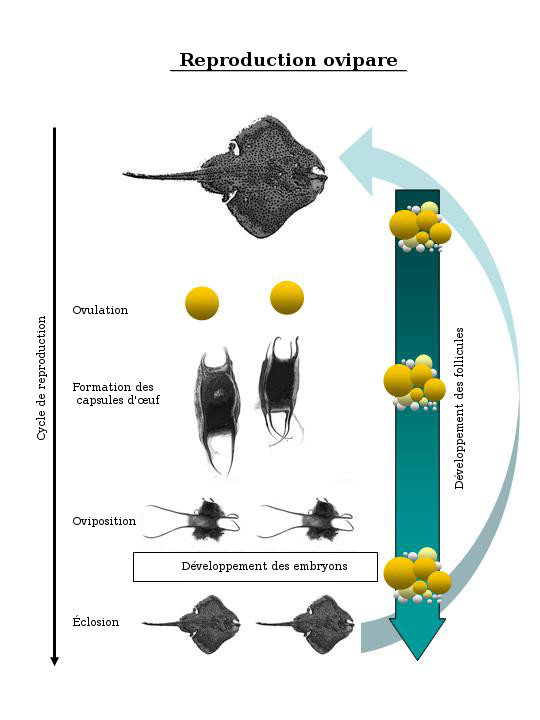
A schematic showing the oviparous reproductive cycle typical of skates., which exhibit a "conveyer belt" type of reproduction. Follicle development occurs in synchrony with oviposition, and a graded series of paired follicles are present at all time within the ovaries.
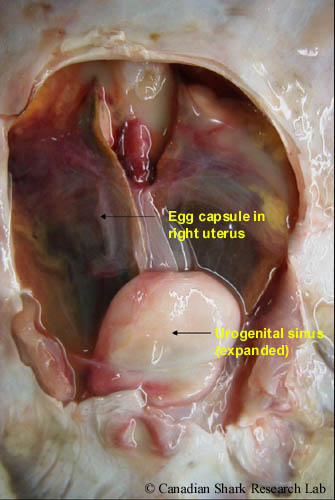
The reproductive tract of a spawning-stage female thorny skate (Amblyraja radiata). Two fully-formed egg capsules are visible in the uteri posterior to the shell glands. The urogenital sinus is greatly expanded.
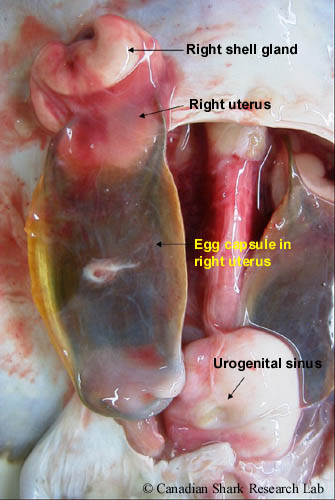
The reproductive tract of a spawning-stage female thorny skate (Amblyraja radiata). The right reproductive tract has been partially removed from the body cavity to show the right shell gland and the fully-formed egg capsule within the uterus.
For a closer look at oviposited eggs, go to the Egg Capsules section of this website. If you want more information on how maturity and reproductive cycles are examined in skates, go to the Skate Research section of this website.
- Date modified: