
External anatomy
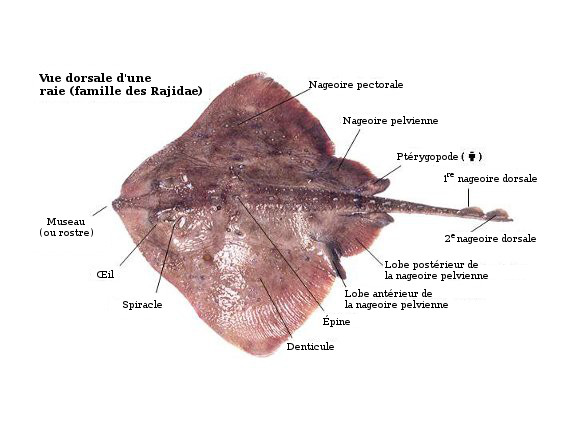
Dorsal view of skate, highlighting important anatomical points.
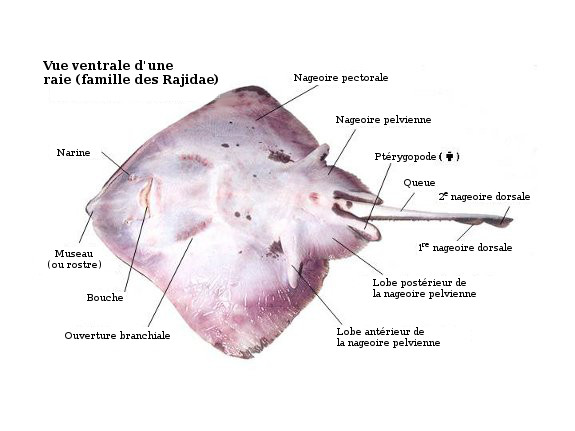
Ventral view of skate, highlighting important anatomical points.
Elasmobranchs have many keen senses that are mostly geared towards helping them find their targeted prey. Which senses are more important to any given species depends largely on the environment in which that species lives. For example, the blind electric ray (Typhlonarke aysoni) which lives at great depths (200 - 900 m) has eyes that are virtually useless, and instead uses electroreceptors to "see". In general, sharks, skates, and rays use the following senses for locating and capturing prey:
- smell (or chemoreception)
- vision (or visual perception)
- hearing (or auditory perception)
- mechanoreception
- electroreception.
Mechanical disturbances picked up by the sensory cells of the lateral line system include those created by moving water, and by sound, vibration, and pressure changes. The Ampullae of Lorenzini detect electrical pulses in the water, and may also help sharks and rays navigate.
Ampullae of Lorenzini
Ampullae of Lorenzini
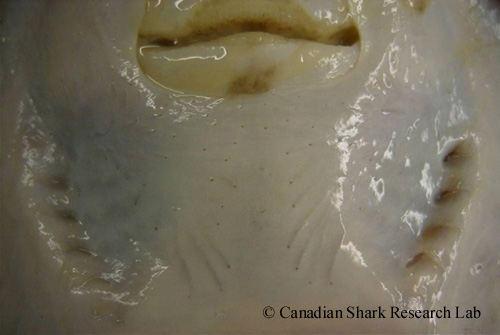
Ampullae of Lorenzini of an immature female winter skate (Leucoraja ocellata), visible on the surface of the skin as small black pores.

Ampullae of Lorenzini of an immature female winter skate (Leucoraja ocellata), visible on the surface of the skin as small black pores.
Unlike teleosts (or bony fishes), which tend to rely heavily on vision, skates and rays have a number of extremely well-developed sensory systems which allow them locate prey and navigate about their environments very efficiently. One of the most interesting sensory adaptations possessed by all elasmobranchs is a complex of small, subcutaneous vesicles located around the head collectively referred to as the ampullae of Lorenzini.
Each ampullae is visible on the surface of the skin as a small pore, with each pore connecting to a long, jelly-filled canal in which a bundle of sensory cells innervated by nerve fibers is located. The ampullae of Lorenzini allow sharks, skates, and rays to detect weak electic fields emitted by fish in distress, and may also respond to mechanical stimuli. In addition, these structures make it possible for benthic feeders - such as skates - to find prey buried in the sand, even in the absence of visual or olfactory cues. Finally, the ampullae may also play an important role in intraspecific communication, particularly among skates and rays, by picking up the weak electrical signals generated by the electric organs of certain species and whose function is otherwise unknown.
Oceanic currents moving in the magnetic field of the Earth produce electric fields that are of the same order of magnitude as the electric fields that sharks and rays are capable of sensing. It has thus been speculated that sharks and rays may use their Ampullae of Lorenzini to orient to the electric fields of oceanic currents. Additionally, the sharks and rays themselves - like all living creatures - produce an electrical field during muscle contractions; as such, the electric field they induce while swimming in the magnetic field of the earth may enable them to sense their magnetic heading.
Electric Organs
Electric Organs

Live winter skate (Leucoraja ocellata), little skate (Leucoraja erinacea), and smooth skate (Malacoraja senta) onboard ship in a holding pen.
Among cartilaginous fishes, electric organs are found only in skates and electric rays. The paired electric organ of electric rays occupies an extensive portion of the pectoral fin. It is very strong, and is used both offensively to capture prey and defensively to discourage predators. In skates, the electric organ is also a paired structure, but it is embedded within the lateral musculature of the tail on either side of the vertebral column. It discharges long-lasting electrical signals thought to be too weak for predatory or defensive purposes. Instead, these signals are thought to play an important role in communicative behavior, such as during reproduction and territoriality. This theory is supported by observations of seasonal variation in the electric organ discharge (EOD) of some species, such as the thornback ray (Raja clavata). In addition, a study on the EOD of the winter skate (Leucoraja oceallata) and the little skate (Leucoraja erinacea) found that EODs occurred most often during behavioural encounters between two or more skates. However, the exact functional role of the electric organ of skates, unlike that of electric rays, is still uncertain.
The tail of skates is often kept very stiff and straight during all activities including swimming, indicating that its primary purpose is not locomotion. Instead, it is though that a rigid tail facilitates the transmission of electrical pulses, which in turn are used for intraspecific communication during important activities such as mating.
Eyes
Eyes

A live male thorny skate (Amblyraja radiata) on deck, with the left eye and spiracle clearly visible
The elasmobranch eye is laterally compressed compared to the basic vertebrate eye, and in addition to the basic structures, possesses a tapetum lucidum. The tapetum is located behind the retina, and is a mirrorlike reflecting layer consisting of parallel, platelike cells filled with crystals. These crystals make the elasmobranch eye extremely efficient in low-light conditions by collectively reflecting light that has already passed through the retina, and re-stimulating the retina a second time. In fact, the elasmobranch eye is twice as effective as that of a cat, and is thought to be one of the most efficient eyes of the animal kingdom.
The eyes of all electric rays, however, are reduced in size and a number of rays are blind. It is thought that these species navigate using electricity instead, which they generate from electric organs in their heads.
Fins
Fins
Sharks, skates and rays have a number of fins which are used for stabilizing, steering, and propulsion. All sharks have one or two fins along the middorsal line called the first and second dorsal fin, both of which act as anti-roll stabilizers. In skates, the dorsal fins are prominent but small and located near the end of the tail, while in rays, the dorsal fins are either absent or greatly reduced in size. Sharks also have a pair of large pectoral fins originating behind the head and extending outward from the trunk that are used for steering. In skates and rays, these fins are enlarged ad "wing-like", often attaching to the side of the head, and are used for propulsion. Finally, sharks, skates and rays have small pelvic fins found on either side of the cloaca which help to maintain stability. In males, the posterior pelvic fins are modified into stout, tube-like copulatory organs called claspers.

A diagram of a typical shark showing the different types of fins. Photo modified from Sharks. 1998. L Compagno, C Simpfendorfer, JE McCosker, K Holland, C Lowe, B Wetherbee, A Bush and C Meyer. Readers Digest Series, Weldon Owen Pty Ltd, Pleasantville NY.
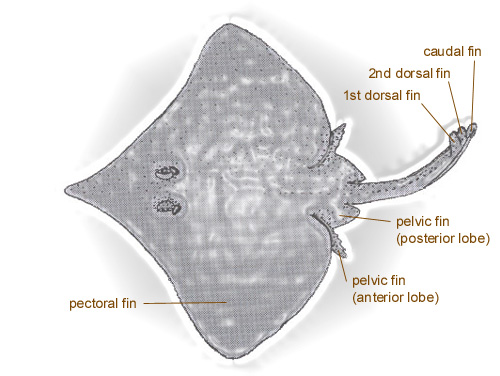
A diagram of a typical skate (family Rajidae) showing the different types of fins. Photo modified from Sharks, Skates and Rays: The Biology of Elasmobranch Fishes. 1999. Edited by William C. Hamlett. John Hopkins University Press, Baltimore, Maryland.
Unlike most other elasmobranchs which propel themselves through the water column using their powerful tails, most batoids (skates and rays) have long, slender tails and therefore must swim by moving their enlarged pectoral fins. Specifically, two types of propulsion are exhibited by batoids: undulatory and oscillatory propulsion. Batoids that exhibit undulatory propulsion swim by passing undulatory waves down their pectoral fins from anterior to posterior, whereas those that exhibit oscillatory appendage propulsion swim by flapping their pectoral fins up and down like the wings of a bird. Most batoids are undulatory appendage propulsors, including all species of skate. Examples of oscillatory appendage propulsors include the eagle and manta rays. The only batoids that locomote using their tails are the electric rays, which locomote by undulating their relatively thick tails.
A spotted eagle ray (Aetobatus narinari). Unlike all other batoids, eagle and manta rays exhibit oscillatory appendage propulsion.
In addition to undulatory propulsion, when maneuvering along the bottom, certain species of skate such as the little skate (Leucoraja erinacea) use an alternate and novel mode of locomotion termed "punting", which is very similar to walking in tetrapods. In these species, the pelvic fins are partially separated into anterior and posterior lobes. The anterior lobe is highly modified, and consists of a functionally distinct leg-like appendage (called the crus) which consists of three flexible joints that allow the skate to push (or "punt") off the substrate. The skate then glides a short distance over the bottom as the crura are repositioned for the next thrust. Asynchronous movement of the crura also allows the skate to turn sharply and quickly to capture prey, a movement that is not possible with pectoral-fin locomotion. Finally, by eliminating mechanical stimulation of the electroreceptors in the pectoral fin regions, punting (or pelvic fin locomotion) may be advantageous when attempting to electrosense potential prey. As such, this novel mode of locomotion is suited to the benthic lifestyle of skates, and is the dominant mode over sandy and muddy bottoms, regardless of the distances traveled.

A spotted eagle ray (Aetobatus narinari).

A spotted eagle ray (Aetobatus narinari).

Ventral surface of an immature little skate (Leucoraja erinacea), with the red arrow demarking the right crus (or modified anterior lobe of the pelvic fin).
Gill Slits
Gill Slits

Ventral side of a mature female thorny skate (Amblyraja radiata), with the red arrow demarking the external gill openings.
The gill slits of a fish are the external openings to the gills. When a fish breathes, water enters through the mouth (or spiracles), passes into the pharynx and through the gill filaments where the respiratory exchange of oxygen and carbon dioxide occurs, and then exits through the gill slits. In sharks, skates and rays, four to seven pairs of gill slits are present on either side of the head.
Nostrils
Nostrils

Ventral side of a mature female thorny skate (Amblyraja radiata), with the red arrows demarking the external nares.
As with sharks, the nostrils (or external nares) of skates and rays are located on the ventral surface of the snout and are the openings to the olfactory organs. In some species, the nostrils may be partially covered by a nasal flap.
Scales
Scales
All cartilaginous fishes have placoid scales on the outside of their bodies, with the number and form of the scales varying widely from one species to the next. In sharks, tiny, overlapping placoid scales are imbedded in the skin and cover most of the body, giving it a sandpaper-like feel. In skates and rays, placoid scales are usually found only as a row or rows of predominant denticles (or thorns) along the midline of the back. In some species, however, placoid scales have been modified into conspicuous structures such as the stinging barbs of stingrays and the defensive spines and thorns that characterize many skate species. The teeth of sharks, skates and rays are also modified placoid scales.
The thorny skate (Amblyraja radiata) - along with other members of the genus Amblyraja - uses its thorns in a novel defensive posture sometimes termed "hedgehog". Like many skates, the thorny skate possesses a median row of bucklers (enlarged dermal denticles) that extend from the scapular region to near the tip of the tail. When startled, the skate folds both its pectoral fins and tail towards its ventral surface and curls into a ball, thereby exposing its heavily thorn-covered dorsal surface and buckler-studded tail to potential predators. Curling into a ball likely protects the thin-skinned belly while also preventing predators from grasping the edge of the pectoral disc, and presentation of the large, sharp dermal denticles may discourage biting by would-be predators.
In addition to defensive spines, mature male skates also have a number of patches of distinct spines on their bodies that serve as anchoring structures during copulation. Lateral to the eye and spiracle are a number of prominent, posteriorly directed dorsal spines that grow in size and number as the skate matures. Similarly, rows of sharp alar spines develop on the outer margins of the pectorals, all of which serve to anchor the male firmly against the body and pelvic fin of the female.

Immature female thorny skate (Amblyraja radiata) with prominent row of thorns visible along midline.

Mature male thorny skates (Amblyraja radiata) showing the defensive "hedgehog" posture.
Spiracles
Spiracles

A mature shorttail (or Jensen's) skate (Amblyraja jenseni), with the red arrow demarking the left spiracle
Skates and rays respire primarily using their spiracles. Spiracles are small, round openings, located on top of and on either side of the head immediately posterior to the eye and anterior to the principal gill opening. These structures can be used in place of the mouth to actively draw water into the gill chambers. This method of respiration is favored by skates and rays because it allows them to breath while lying motionless on the bottom waiting to ambush prey, or buried within the bottom substrate to avoid predators.
Tail
Tail
The tail (or caudal) fin of skates and rays plays a different role than in most other elasmobranchs. In sharks, the caudal fin is a distinct heterocercal shape and is the major source of propulsion. Side to side movement of the tail in a sculling motion is used to generate the forward thrust that pushes the shark through the water. In comparison, the tails of rays tends to be long and slender and are not used for propulsion, with the exception being electric rays. In stingrays and batrays, the tail contains a spine modified from a placoid scale with a venom gland at its base. The spine is whipped about by the tail in defense against large predators such as sharks.

A tropical species, the southern stingray (Dasyatis americana) (shown above) has a long slender tail typical of stingrays and batrays.

A southern stingray (Dasyatis americana). Note the long, slender tail with a venomous spine.

A spotted eagle ray (Aetobatus narinari). Note the very long, slender tail with a venomous spine

Live skates in a holding tank at the Bedford Institute of Oceanography. Note the rigid, muscular tails.

An embryonic winter skate (Leucoraja ocellata) removed from its egg capsule. Note the extended tip of the tail
The significance of the tail in adult skates is less obvious, since it does not appear to be involved in locomotion or active defense. During most behaviors (including swimming), it is maintained in a rigid state. However, it is possible that the persistence of prominent muscular tail in skates is related, at least in part, to the presence of the electric organ, with tail rigidity aiding the spread of current during electric organ discharge.
In addition, during embryonic development in skates, the tail plays an important role is circulating seawater through the egg case. The elongated tip of the tail (visible in the photo to the right) which aids greatly in circulating the seawater is lost soon after hatching.
Teeth
Teeth

Ventral view of the jaws of a mature male shorttail (or Jensen's) skate (Amblyraja jenseni)
The teeth of elasmobranchs are derived from dermal denticles (or placoid scales) and are thus structurally similar. Each tooth consists of a root that is firmly implanted in the connective tissue of the jaw and a visible crown - often with pointed projections called cusps - that protrudes from the jaw. Most elasmobranchs are specialized predators, with their dentition reflecting both the manner in which they feed and their preferred diet. Typically, elasmobranch teeth are designed for seizing, cutting or grasping fast-moving prey. The teeth of skates and rays, on the other hand, are flattened and pavement-like with low, blunt crowns perfect for crushing their hard-shelled invertebrate prey.
One aspect of the dentition of elasmobranchs that is shared among all species is the continual shedding and replacement of teeth. A close examination of the jaws of a skate (for example) reveals the front biting row, followed by row upon row of teeth ready to replace any outer teeth that are lost through wear and tear.
Interestingly, in many species of elasmobranch, the teeth of adult males are different in shape from those of females and immature males. This is referred to as sexual heterodonty or dental sexual dimorphism, and is often confined to the anterior teeth. In sharks, and particularly in batoids (such as the Atlantic stingray, Dasyatis sabina), sexual heterodonty appears to function not in feeding, but in courtship by allowing mature males to firmly grasp on to the females with their mouths. Many northwest Atlantic skates exhibit dental sexual dimorphism, including the winter skate, the little skate, the barndoor skate, the smooth skate, and the thorny skate. In these species, the teeth of both sexes are blunt or rounded until individuals reach sexual maturity. Just prior to maturity, the teeth of the males develop sharp conical cusps and the shape of the mouth becomes more sinuous. In the Atlantic stingray, there is even a periodic shift in male dentition over the reproductive season, with male teeth becoming more cuspidate and less molariform prior to mating.
- Date modified: