
Haplosporidian Parasites of Abalone
On this page
Category
Category 3 (Host Not in Canada)
Common, generally accepted names of the organism or disease agent
- Haplosporidian parasite of New Zealand paua.
- Haplosporidian parasite of European abalone.
Scientific name or taxonomic affiliation
According to Hine et al. (2009) the taxonomy of haplosporidians needs a thorough revision. Nevertheless, two different haplosporidians have been reported from abalone. All information pertaining to each of these parasites was assigned a letter code which was consistently applied throughout this page.
- This parasite was not specifically identified because the features necessary for a correct description of the genus and species of organisms in the phylum Haplosporidia were not characterized. The characterization of this parasite was not possible because no sporulated stages nor spores were observed. Transmission electron microscopy and molecular studies indicate that the parasite is a novel species in perhaps a new genus at the base of the phylum Haplosporidia and ancestral to other genera in this group (i.e., Urosporidium, Haplosporidium, Bonamia, and Minchinia) (Reece et al. 2004). Burreson and Ford (2004) confirmed that this parasite was correctly assigned to the phylum Haplosporidia.
- Haplosporidium montforti (Azevedo et al. 2006).
Geographic distribution
- A commercial abalone culture facility in New Zealand (Diggles et al. 2002, Hine et al. 2002).
- Detected in Haliotis tuberculata imported from Ireland and experimentally grown in Galicia (NW Spain). Although H. montforti was not detected by light microscopy in juvenile abalone imported directly from Ireland, molecular analyses indicated that this parasite was present in juvenile abalone from Ireland before their introduction into Galician waters (Balseiro et al. 2006). Molecular analyses also indicated that juvenile H. tuberculata imported from France were infected. However, H. montforti DNA was not detected in a small sample (n=29) of indigenous (natural) populations of adult H. tuberculata from Galician waters (Ría de Arousa) close to the abalone culture locations (Balseiro et al. 2006).
Host species
- Haliotis iris.
- Haliotis tuberculata.
Impact on the host
- Moribund H. iris from a population of juveniles experiencing mortalities (peak of 24% per week with a total mortality approaching 90% within 6 months) at a commercial abalone culture facility had heavy systemic infections of a uni- to multi-nucleate protist. The parasite was confirmed by transmission electron microscopy and molecular studies to be a haplosporidian. Mortalities associated with the infection were most severe during the summer and early autumn when water temperatures peaked at 21 °C. Results of experimental exposure in the laboratory indicated that the parasite was not transmitted between H. iris by cohabitation (for 3 months) nor injection (Diggles et al. 2002). A subsequent survey of 1094 H. iris from 5 spat production farms and 3 grow-out farms in New Zealand did not detect this parasite (Diggles and Oliver 2005).
- Haliotis tuberculata originally from Ireland and experimentally grown in plastic barrels suspended from rafts in Galicia (NW Spain) experienced high mortalities linked to Haplosporidium montforti. All dead abalone were heavily parasitized by H. montforti (Azevedo et al. 2006). However, the presence of a proteobacteria "Candidatus Xenohaliotis californiensis" , a pathogen from the family Rickettsiaceae, in the gastrointestinal epithelial cells of moribund abalone may have caused the disease to be more extreme (Balseiro et al. 2006).
Diagnostic techniques
Gross Observations
- Haliotis iris from raceways with increased mortalities exhibit behavioural abnormalities including lethargy, loss of righting reflex and were easily detached from the surface. Apart for these behavioural abnormalities that are not specific to infection with this parasite and oedema and pale lesions in the foot and mantle, no reliable gross signs of disease were noted.
- Signs of disease in H. tuberculata included dark foot pigmentation, loss of surface adherence and a limited ability to right themselves after they were set on their backs (Azevedo et al. 2006, Balseiro et al. 2006). However, these signs are not specific to infection with this parasite and they cannot be used as a tool to discrimination among diseases of abalone (Balseiro et al. 2006).
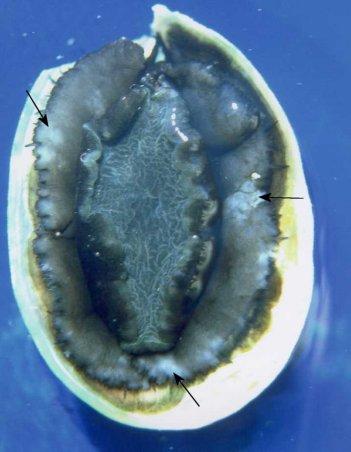
Figure 1. Gross appearance of a juvenile Haliotis iris from a commercial culture facility heavily infected with a haplosporidian parasite. Note the blotchy appearance of the epipodium (arrows). Image provided by Ben Diggles Ph. D., DigsFish Services.
Wet Mounts
- Multinucleate plasmodia up to 25 µm in diameter with up to 17 nuclei were detected in wet preparations of haemolymph.
- Refringent sporonts and free spores with an operculum are evident in tissue squashes of fresh, heavily infected abalone (Balseiro et al. 2006). Free mature spores of H. montforti viewed via differential interference contrast light microscopy (Nomarski-DIC) showed that the spores were ornamented by 4 filaments (20-28 µm in length), some of which seemed branched. Two of the filaments were attached opposite each other at the apical zone beneath the operculum and the other two were also opposite each other but attached at the basal portion of the spore (Azevedo et al. 2006).
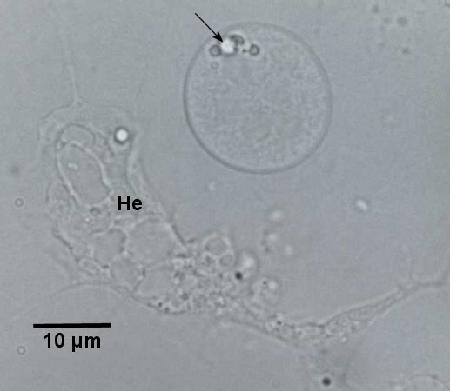
Figure 2. Unstained hemolymph preparation from infected Haliotis iris showing spherical haplosporidian plasmodium with prominent lipid droplets (arrow) and 6 nuclei above a typical irregularly shaped haemocyte (He) adhered to the glass slide. Image provided by Ben Diggles Ph. D., DigsFish Services.
Smears
- Up to 17 nuclei in one plasmodium were evident in air-dried haemolymph smears stained with a Giemsa type stain (Diggles et al. 2002).
- Smear of connective tissue from the foot of heavily infected abalone showed various developmental stages and spores of H. montforti (Azevedo et al. 2006).
Histology
- Systemic infection with uni- to multi-nucleate haplosporidian-like plasmodia (ranging is size from 3.5-8.9 x 3.5-16.7 µm with an average of about 5.5 x 6.9 µm) in cultured juvenile H. iris. Up to 9 spherical nuclei, each with emarginated chromatin and about 2.3 µm in diameter, were observed within cross sections of a single plasmodia but on average only about 2 nuclei were observed in each plasmodium. Each individual plasmodium was usually surrounded by a clear area, which facilitated detection. These distinctive halos may be evidence of extra cellular toxic products produced by the plasmodia or differential shrinkage of the plasmodia during fixation. Early stages of infection were characterised by a small number of plasmodia in the connective tissues surrounding the gut, amongst glial cells adjacent to nerves in the mantle and foot and within gill lamellae (Diggles et al. 2002).
Spore stages of the parasite have not been described.
Although haemocytes were sometimes associated with the plasmodia in light infections, relatively few or no host haemocytes were visible in heavily infected H. iris (even in comparison to apparently healthy uninfected paua). In heavy infections, large numbers of average sized plasmodia were present in the haemolymph sinuses of all organs and within the tissues of the gills, heart, kidneys, mantle, epipodium of the foot, and connective tissue of the digestive gland (Diggles 2002).
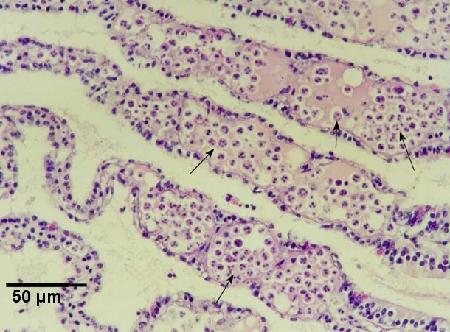
Figure 3. Large numbers of haplosporidian plasmodia (arrows) in the haemal sinuses of the gills of a heavily infected Haliotis iris.
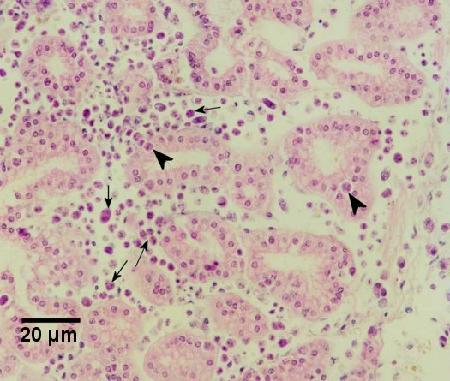
Figure 4. Haplosporidian plasmodia in the connective tissues (arrows) and occasionally in the tubule epithelium (arrowheads) of the right kidney of a heavily infected Haliotis iris.
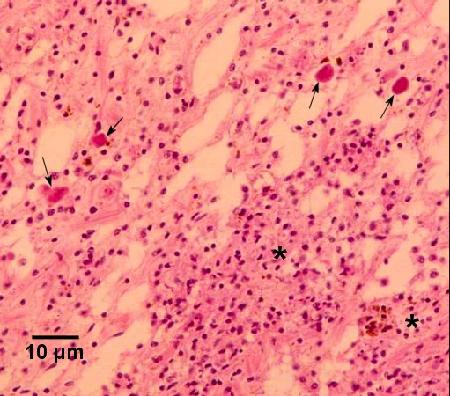
Figure 5. Haplosporidian plasmodia (arrows) in the connective tissue adjacent to the gut and associated with multifocal haemocytosis (*) of a moderately infected Haliotis iris. Images in Figs. 3 to 5 provided by Ben Diggles Ph. D., DigsFish Services.
- Various developmental stages of H. montforti were observed in the connective tissue of the digestive gland, kidney, mantle, gills and foot muscle. Round uninucleated plasmodia (3.2 ± 0.2 µm in diameter with nuclei of 1.3 ± 0.1 µm) observed in the connective tissue of gills and digestive tract may be the initial stages of the infection. Initial developmental stages included plasmodia with one to four nuclei and eventually multinucleated plasmodia with up to eight nuclei were found in connective tissue close to the digestive epithelium. Abalone with advanced stages of infection had plasmodia (often round, surrounded by an irregular membrane and about 15.3 µm in diameter) undergoing karyokinesis and forming sporonts. Each sporont developed into approximately 100 immature spores with basophilic nuclei and cytoplasm surrounded by a thin wall. Immature spores gave rise to refringent mature spores by thickening of the external wall and formation of a typical opercular lid (Balseiro et al. 2006). The H. montforti infection was associated with haemocytic infiltration in foot muscle and connective tissue of the digestive tract and gills. In foot muscle, the infection causes a reduction in muscle fibers, loss of muscle bundle orientation and foot muscle atrophy. Heavy infections eventually caused total destruction of the architecture of all organs (Balseiro et al. 2006). In heavily infected abalone, there was a nearly complete substitution of host tissue by developmental stages of the parasite (Azevedo et al. 2006).
Electron Microscopy
- Ultrastructural features consistent with other haplosporidians that were identified in this parasite include: multinucleate plasmodia, mitochondria with tubular cristae, anastomosing endoplasmic reticulum, multivesicular bodies and haplosporogenesis (haplosporosome-like bodies produced from nuclear membrane-bound Golgi that matured into haplosporosomes). This abalone parasite differs from previously described haplosporidians in the apparent absence of a persistent mitotic spindle and the presence of intracytoplasmic coccoid to rod-shaped bacteria resembling Rickettsiales-like prokaryotes (Hine et al. 2002).
- Systemic infections of H. montforti had uninucleated and multinucleated plasmodia containing spherical nuclei. Internally, the uninucleated endosporoplasm contained typical haplosporidian structures, such as; haplosporosomes, a spherulosome, and mitochondria with vesicular cristae. The plasmodia occurred among sporocysts containing different spore maturation stages (Azevedo et al. 2006). The nucleus of the uninucleated plasmodia divided successively, giving rise to multinucleate plasmodia, containing from 100 to 120 nuclei. Later, the plasmodia developed into sporonts inside sporocysts with irregular contours. Each of their nuclei gave rise to uninucleate sporoblasts. At the next phase of development, a very irregular membranous group of cisternae began to differentiate in the cytoplasm of each sporoblast, surrounding each nucleus and the adjacent cytoplasm. Each sporoblast differentiated into a spore. This process was characterized by the appearance of dense blisters of amorphous material at the periphery that gradually formed the prespore wall and pre-operculum. Simultaneously, in the endosporoplasm, the spherulosome and several haplosporosomes were formed. During the final phase of the maturation process, the spores became gradually denser, and the endosporoplasmic structures were barely visible (Azevedo et al. 2008). The ultrastructural morphology of the spores and their surrounding ornaments attached to the spore wall was described from scanning and transmission electron microscopy. The spores were spherical to slightly ellipsoidal (2.42 ± 0.5 by 2.31 ± 0.6 µm). The apical zone of the spore wall was modified into a complex opercular system covering a circular orifice that measured about 0.5 µm in diameter. The operculum was connected to the spore wall by a hinge. The spore wall was about 110 nm thick and consisted of three layers: an electron-dense proteinaceous outer layer about 50 nm thick; an electron-light middle layer about 20 thick; and an inner electron–dense layer about 40 nm thick that was less dense than the outer layer. Four filaments (20 nm to 28 µm long) attached to the cell wall were composed of the same material that formed the wall. The cross-sections through the base of these filaments showed L-like, T-like and X-like sections and the apical region of the filaments were circular and ellipsoidal in cross-sections (Azevedo et al. 2006).
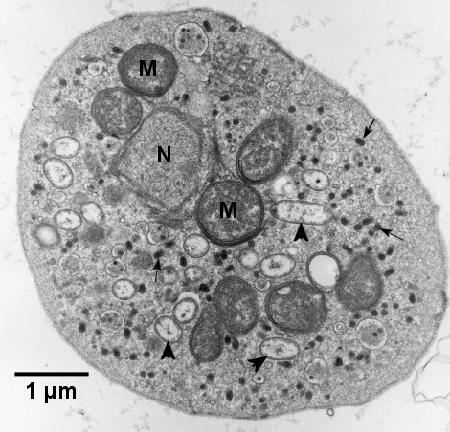
Figure 6. Electron micrograph of a haplosporidian plasmodium from Haliotis iris containing a section through the edge of one nucleus (N), mitochondria (M), haplosporosomes (arrows) and Rickettsiales-like organisms (arrowheads). Image provided by Ben Diggles Ph. D., DigsFish Services.
DNA Probes
- The in situ hybridisation assay designed to detect rDNA of Bonamia ostreae (Cochennec et al. 2000) gave a moderately strong reaction with the paua haplosporidian and demonstrated the presence of low numbers of plasmodia in the connective tissue adjacent to the gut and amongst glial cells around the periphery of the nerves in lightly infected H. iris (Diggles et al. 2002). A fragment of the small-subunit rDNA gene has been sequenced (1967 bp in length after removal of the primer sequences) and the sequence deposited into GenBank (accession number AF492442). This sequence was used to determine the taxonomic affiliations of the parasite (Reece and Stokes 2003).
- The molecular characterization of the small subunit ribosomal RNA (SSU rRNA) gene was determined for H. montforti. The 1872-bp sequence was deposited in the GenBank database (Accession No. DQ219484). This sequence was different from previously reported haplosporidian SSU rRNA gene sequences (Azevedo et al. 2006). A nested polymerase chain reaction (PCR) test was designed for the detection of H. montforti using primers developed from the sequence deposited in GenBank (Balseiro et al. 2006).
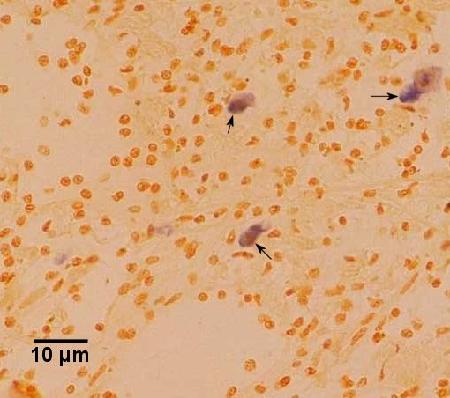
Figure 7. Serial section to that illustrated in Figure 5 but stained by in situ hybridisation with the DNA probe described by Cochennec et al. (2000) indicating a staining reaction with the haplosporidian plasmodia (arrows). Image provided by Ben Diggles Ph. D., DigsFish Services.
Methods of control
a. and b. No known control methods. Research on another species of haplosporidian (Haplosporidium nelsoni, Ford et al. 2001) suggests that passing incoming water through a 1 µm filter followed by UV irradiation may exclude infective stages from the water supply of a commercial tank facility. Surveys of wild stocks of juvenile and adult H. iris and H. tuberculata could also be used to establish the distribution of the parasite in wild stocks to hopefully avoid the inadvertent transfer of the disease.
References
Azevedo, C., P. Balseiro, G. Casal, C. Gestal, R. Aranguren, N.A. Stokes, R.B. Carnegie, B. Novoa, E.M. Burreson and A. Figueras. 2006. Ultrastructural and molecular characterization of Haplosporidium montforti n. sp., parasite of the European abalone Haliotis tuberculata. Journal of Invertebrate Pathology 92: 23-32.
Azevedo, C., G. Casal and J. Montes. 2008. Ultrastructural developmental cycle of Haplosporidium montforti (Phylum Haplosporidia) in its farmed abalone host, Haliotis tuberculata (Gastropoda). Journal of Parasitology 94: 137-142.
Balseiro, P., R. Aranguren, C. Gestal, B. Novoa and A. Figueras. 2006. Candidatus Xenohaliotis californiensis and Haplosporidium montforti associated with mortalities of abalone Haliotis tuberculata cultured in Europe. Aquaculture 258: 63–72.
Cochennec, N., F. LeRoux, F. Berthe and A. Gerard. 2000. Detection of Bonamia ostreae based on small subunit ribosomal probe. Journal of Invertebrate Pathology 76: 26-32.
Diggles, B.K. and M. Oliver. 2005. Diseases of cultured paua (Haliotis iris) in New Zealand. In: Walker, P.J., R.G. Lester, M.G. Bondad-Reantaso (eds.) Diseases in Asian Aquaculture V. Proceedings of the 5th Symposium on Diseases in Asian Aquaculture. Fish Health Section, Asian Fisheries Society, Manila. pp. 275-287.
Diggles, B.K., J. Nichol, P.M. Hine, S. Wakefield, N. Cochennec-Laureau, R.D. Roberts and C.S. Friedman. 2002. Pathology of cultured paua Haliotis iris infected with a novel haplosporidian parasite, with some observations on the course of disease. Diseases of Aquatic Organisms 50: 219-231.
Ford, S.E., Z. Xu and G. Debrosse. 2001. Use of particle filtration and UV irradiation to prevent infection by Haplosporidium nelsoni (MSX) and Perkinsus marinus (Dermo) in hatchery-reared larval and juvenile oysters. Aquaculture 94: 37-49.
Hine, P.M., S. Wakefield, B.K. Diggles, V.L. Webb and E.W. Maas. 2002. Ultrastructure of a haplosporidian containing Rickettsiae, associated with mortalities among cultured paua Haliotis iris. Diseases of Aquatic Organisms 49: 207-219.
Hine, P.M., R.B. Carnegie, E.M. Burreson and M.Y. Engelsma. 2009. Inter-relationships of haplosporidians deduced from ultrastructural studies. Diseases of Aquatic Organisms 83: 247-256.
Reece, K.S. and N.A. Stokes. 2003. Molecular analysis of a haplosporidian parasite from cultured New Zealand abalone Haliotis iris. Diseases of Aquatic Organisms 53: 61-66.
Reece, K.S., M.E. Siddall, N.A. Stokes and E.M. Burreson. 2004. Molecular phylogeny of the haplosporidia based on two independent gene sequences. The Journal of Parasitology 90: 1111-1122.
Citation Information
Bower, S.M. (2014): Synopsis of Infectious Diseases and Parasites of Commercially Exploited Shellfish: Haplosporidian parasites of abalone.
Date last revised: April 2014
Comments to Susan Bower
- Date modified: