
Anatomie internes
Squelette
Squelette
Le squelette interne des raies (ou endosquelette), à l'instar de celui des requins, ne contient aucun os : il est entièrement constitué de cartilage. Le cartilage est une matière résistante et durable plus légère et plus flexible que les os, ce qui empêche les élasmobranches (qui sont dépourvus de vessie natatoire) de rester à flot et leur permet de réaliser des virages plus serrés que les autres poissons. Cependant, certaines parties du squelette des élasmobranches (comme le crâne, les vertèbres et les épines cutanées) sont souvent renforcées par un dépôt de calcium et de sels (processus appelé calcification). Lorsque les structures du squelette des élasmobranches sont suffisamment calcifiées, elles ressemblent parfois à des os et sont aussi résistantes qu'eux, sans toutefois peser autant. Le processus de calcification varie beaucoup d'une espèce à l'autre. Certaines espèces, surtout celles des grandes profondeurs et les formes primitives, affichent une faible calcification tandis que d'autres, comme les requins pélagiques, montrent une très forte calcification.
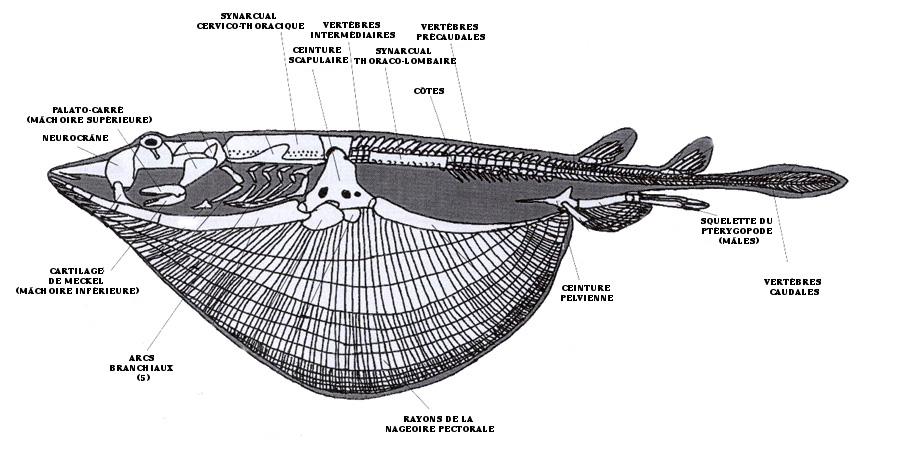
Diagramme de la vue latérale d'un squelette idéalisé de batoïde illustrant les principales structures du squelette. Photo modifiée tirée de Sharks, Skates and Rays: The Biology of Elasmobranch Fishes. 1999. Publié sous la direction de William C. Hamlett. John Hopkins University Press, Baltimore, Maryland.
Les centra vertébraux des élasmobranches sont de forme cylindrique et biconcave, et ils peuvent généralement être classés selon deux principaux types : vertèbre précaudale et vertèbre caudale. Le nombre de vertèbres qu'aura un individu dans sa vie est déterminé durant le développement embryonique, période qui marque également le début de la calcification des vertèbres. Les vertèbres précaudales commencent généralement à se former avant les vertèbres caudales, et toutes les vertèbres semblent croître tout au long de la vie des requins et des raies. Cela permet de déterminer l'âge des individus au moyen des centra vertébraux. Chez les élasmobranches, on détermine l'âge d'un individu en comptant les anneaux vertébraux qui se forment chaque année, un peu comme l'écorce des arbres. Cette méthode est analogue à l'utilisation des otolithes (ou « pierres de l'oreille ») pour déterminer l'âge des poissons osseux. Pour en apprendre davantage sur la façon dont on détermine l'âge des raies, veuillez consulter la section Recherche sur les raies du site Web.
Chez les batoïdes, la colonne vertébrale est plus élaborée que chez les requins. Plus particulièrement, de derrière le neurocrâne (le crâne des élasmobranches) jusqu'à l'extrémité antérieure de la colonne vertébrale, les vertèbres précaudales sont intégrées dans un synarcual cervico-thoracique, qui peut comprendre entre 4 et 45 vertèbres selon l'espèce (figure 1). Cette structure rigide est beaucoup moins flexible que le reste de la colonne vertébrale, et elle sert à supporter les très longues nageoires pectorales. Certaines espèces de myliobatidés possèdent même un deuxième synarcual!

Vue ventrale d'un embryon de raie épineuse (Amblyraja radiata) sous une lumière reflétée. On peut clairement voir les rayons endosquelettiques de la nageoire pectorale allant jusqu'aux bords distaux de la nageoire pectorale de droite.
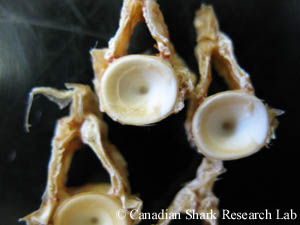
Vertèbre thoracique d'une raie épineuse (Amblyraja radiata).

Centra vertébral d'un grand requin-marteau (Sphyrna mokarran). Les anneaux de croissance sont clairement visibles.
Foie
Foie

Raie épineuse (Amblyraja radiata) mature femelle disséquée pour montrer le gros foie lobé (qui a été retiré de la cavité abdominale). Le foie est le premier organe que l'on trouve lorsque l'on incise le ventre pour exposer la cavité abdominale.
Le foie des élasmobranches est un gros organe lisse et très huileux qui occupe presque toute la cavité abdominale. Il peut représenter jusqu'à 25 % du poids du poisson. Lorsque l'on incise le ventre depuis l'axe des nageoires pelviennes jusqu'à l'extrémité antérieure de la cavité abdominale, c'est le foie que l'on trouve en premier. Il présente deux gros lobes pointus, dont la couleur varie entre le gris-vert et le brun-rouge foncé. Le foie des requins et des raies a deux fonctions. La première, comme chez tous les animaux, est de concentrer les réserves en lipides et donc d'emmagasiner l'énergie. La deuxième fonction du foie des élasmobranches consiste à servir d'organe hydrostatique en emmagasinant des huiles plus légères que l'eau (à densité moindre). Ces huiles contrebalancent la tendance des élasmobranches à couler en diminuant la densité et en augmentant la flottabilité de l'animal dans son ensemble, de sorte que l'animal atteint un état de flottabilité neutre. Sans un tel foie, les requins et les raies auraient du mal à s'éloigner du fond, car ils ne possèdent pas de vessie natatoire qui caractérise les poissons osseux.
Système digestif
Système digestif
La deuxième structure la plus flagrante dans la cavité abdominale des requins et des raies est le système digestif, qui est constitué de deux organes voisins : l'œsophage et l'estomac. L'extrémité antérieure de l'estomac (aussi appelée estomac cardiaque) est un sac en forme de « J » qui est rattaché à la partie postérieure de l'estomac (appelée estomac pylorique) et qui est courbé antérieurement. L'estomac pylorique se termine à une constriction appelée le pylore, qui mène au court duodénum puis à la valvule spirale de l'intestin, plus grosse et tout enroulée et tordue. La fonction de la valvule spirale de l'intestin est d'accroître la superficie pour la digestion et l'absorption des aliments, tout en laissant assez d'espace dans la cavité abdominale pour le gros foie et les embryons en développement (dans le cas des raies de l'ordre des Myliobatiformes) ou les capsules d'œuf (dans le cas des raies de l'ordre des Rajiformes). La valvule spirale, quant à elle, mène au rectum puis à l'anus, qui s'ouvre dans le cloaque, une cavité qui sert d'orifice pour les voies intestinales, urinaires et génitales.
Pancréas
Pancréas
Le pancréas est une glande qui favorise la digestion en sécrétant des enzymes digestifs dans le duodénum. Il est constitué de deux lobes reliés : un lobe ventral, qui contient un canal par lequel les sécrétions pancréatiques entrent dans le duodénum, et un lobe dorsal. Les deux lobes sont généralement de couleur rosée.
Rate
Rate
La rate est un organe brun foncé, de forme triangulaire ou légèrement allongée, qui repose contre l'estomac. Toutefois, elle ne joue pas de rôle dans le processus digestif. Elle fait plutôt partie du système lymphatique, qui constitue un élément essentiel du système immunitaire.
Glande rectale
Glande rectale
La glande rectale est un petit organe en forme de doigt qui emmagasine d'importantes quantités de sel excédentaire (ou chlorure de sodium) du sang en vue de les excréter par l'anus. Plus précisément, elle secrète une solution incolore dans le rectum au moyen d'un petit canal, solution dont la teneur en chlorure de sodium est le double de celle contenue dans le plasma sanguin. Cet organe est très important pour les élasmobranches, dont le foie produit une grande quantité d'urée, rendant ainsi ces poissons marins légèrement hyperosmotiques dans l'eau de mer (c.-à-d. que leur concentration interne de solutés est plus élevée que celle de l'eau de mer dans laquelle ils se trouvent, de sorte qu'ils absorbent constamment de l'eau et du sel).
Reins
Reins
Les reins font partie du système urogénital et jouent un rôle dans la fabrication et le transport de l'urine, ainsi que dans la régulation de la teneur plasmatique en ruée. Chez les raies, les reins ont la forme d'une demi-lune ou d'un ruban, sont aplatis dorsoventralement et sont rouge foncé. Ils sont très lobés et reposent sur le dos contre la colonne vertébrale, à l'extérieur de la cavité abdominale. Une membrane épaisse connue sous le nom de péritoine sépare les reins du reste de la cavité abdominale. Les reins sont drainés dans le cloaque par les urètres.
Système reproducteur mâle
Système reproducteur mâle

Raie à queue de velours (Malacoraja senta) mâle immature ouverte afin de révéler les testicules blanc-jaune aux lobes indistincts. Noter les petits ptérygopodes qui ne sont pas encore développés.
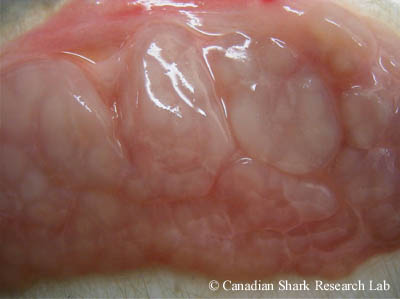
Les organes du système urogénital du mâle comprennent les gonades mâles (ou testicules), les canaux génitaux, la glande de Leydig et la glande alcaline. Lorsqu'on procède à l'ablation du système digestif, on peut voir la paire de testicules plats, de couleur blanchâtre ou jaunâtre, qui sont situés à l'extrémité antérieure de la cavité abdominale, en arrière du foie. Chez de nombreuses espèces, les testicules sont enveloppés de ce que l'on appelle l'organe épigonal, qui semble jouer un rôle dans la formation des cellules sanguines. Chaque testicule est composé d'un regroupement de lobes de formes irrégulières; chez les requins immatures, les testicules sont une masse discrète de tissu blanchâtre aux lobes peu visibles, tandis que chez les requins matures, les lobes sont distincts et bombés.
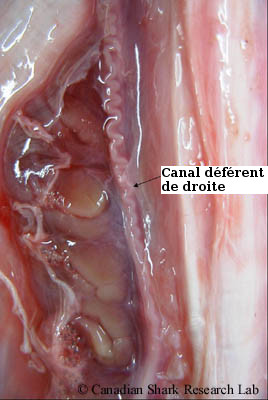
Les spermatozoïdes (ou spermatocytes) sont produits et se développent dans les lobes de chaque testicule (processus appelé spermatogénèse). Les spermatozoïdes matures sont ensuite transportés dans de minuscules canaux (appelés canaux efférents) jusque dans un épididyme torsadé, que l'on peut apercevoir le long de la colonne vertébrale, de chaque côté de l'aorte dorsale. Le sperme continue son chemin vers la partie postérieure de l'animal, en passant de l'épididyme au ductus deferens (ou canal déférent). C'est là que le sperme est stocké avant la copulation, et où les spermatophores se forment. Les spermatophores sont de petits sacs ovoïdes ou sphériques comportant un grand nombre de spermatozoïdes. Ils servent à protéger le sperme et à empêcher toute fuite ou perte de sperme dans l'eau durant l'accouplement. L'épididyme et le canal déférent reçoivent une sécrétion visqueuse produite par la glande de Leydig adjacente qui semble stimuler le mouvement des spermatozoïdes.
Chez les mâles immatures, le canal déférent est un mince tube droit, tandis que chez les mâles matures, la partie antérieure du canal déférent est torsadée et sinueuse, couvrant presque tous les reins. On distingue également les mâles immatures des mâles matures par la partie postérieure du canal déférent qui, chez ces derniers, est très élargie pour former les vésicules séminales. Du côté ventral de chaque vésicule séminale se trouve une protubérance (ou dilatation latérale) appelée sac de sperme; les deux sacs de sperme sont reliés plus loin et forment le sinus urogénital, qui se termine en tant que papille urogénitale et se déverse dans le cloaque. En fait, le terme « sac de sperme » ne convient guère, puisque ces structures ne contiennent pas de sperme. Chez diverses espèces de raies, ces structures sont en fait une glande spécialisée (appelée glande alcaline ou de Marshall) contenant un fluide très alcalin qui neutralise l'urine acide et peut aussi jouer un rôle dans la protection du sperme et la formation de bouchons copulatoires. Le cloaque est situé entre les nageoires pelviennes et est présent chez les deux sexes. Il s'agit d'une cavité qui sert d'orifice pour les voies intestinales, urinaires et génitales.



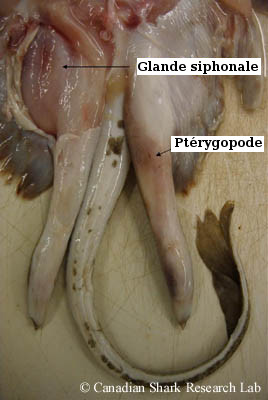
Chez les élasmobranches mâles, la bordure interne des nageoires pelviennes constitue une paire d'appendices copulatoires en forme de tube appelés myxoptérygies, ou plus communément ptérygopodes. Durant l'accouplement, les ptérygopodes sont ramenés vers l'avant : l'un d'entre eux est inséré dans la femelle, et le sperme (ou les spermatophores) est acheminé le long du sillon dorsal longitudinal du ptérygopode jusque dans le cloaque de la femelle. Chez les requins, le sperme est éjecté avec force du cloaque à l'extrémité du sillon du ptérygopode par un organe musculaire contractile appelé sac siphonal. Ce dernier est situé sous la peau du ventre. Plus précisément, avant l'accouplement, de l'eau de mer est pompée dans les sacs par la flexion des ptérygopodes, puis est éjectée lorsque le sac siphonal se contracte, propulsant ainsi le sperme dans la femelle.
Les batoïdes sont dépourvus de sac siphonal; toutefois, ils sont dotés d'une paire de glandes siphonales sur le ventre, qui sont recouvertes d'un muscle strié et se vident dans un petit sac près des nageoires pelviennes. Lorsque le muscle strié se contracte, les sécrétions des glandes siphonales sont propulsées dans le sac des ptérygopodes, qui force ensuite les sécrétions à passer dans le sillon des ptérygopodes, où tout se mélange au sperme du cloaque. Les sécrétions des glandes agissent à titre de lubrifiant et de moyen de transport pour le sperme, et elles scellent le sillon du ptérygopode pour empêcher la dilution de la semence et la perte de sperme dans l'eau de mer. L'accouplement des batoïdes est généralement long, durant environ de 1 à 4 heures; toutefois, il est rare que l'on puisse observer cette activité chez les élasmobranches.
Pour en apprendre davantage sur la façon dont la maturité et les cycles de reproduction sont étudiés chez les raies, veuillez consulter la section Recherche sur les raies du site Web.
Système reproducteur femelle
Système reproducteur femelle
Chez les femelles, l'ablation du système digestif expose le système urogénital, qui est constitué des gonades (ou ovaires), de l'ostium, de l'oviducte, de la glande nidamentaire (ou coquillière), des utérus et du sinus urogénital. Les ovaires, qui sont d'un jaune blanchâtre, sont situés à l'extrémité antérieure de la cavité abdominale, à l'arrière du foie.
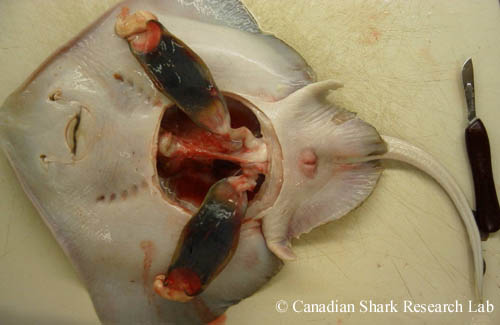
Raie épineuse (Amblyraja radiata) femelle mature ouverte pour examen de son système reproducteur. On peut voir deux capsules d'œuf entièrement formées dans les utérus derrière les glandes nidamentaires.
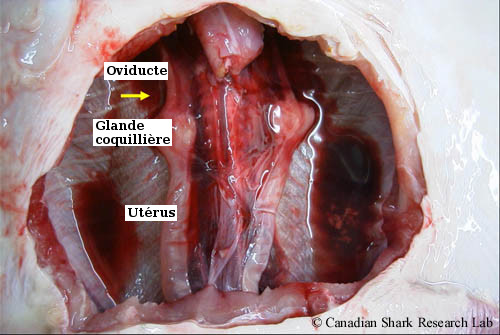
Système reproducteur d'une jeune raie épineuse (Amblyraja radiata) femelle. La flèche jaune indique l'emplacement de l'ovaire droit, qui a été enlevé pour montrer les structures reproductrices dorsales.

Système reproducteur d'une raie épineuse (Amblyraja radiata) femelle mature. Noter les gros ovules vitellogènes dans ses ovaires, les glandes nidamentaires dilatées, les utérus flasques et le sinus urogénital enflé.
Chez les femelles immatures, les ovaires sont lisses et minces. Lorsque la femelle atteint sa maturité, de nombreux petits oocytes blancs deviennent visibles dans le tissu conjonctif des ovaires, ce qui donne à ces derniers une apparence granulée. Chez les femelles qui ont atteint la maturité sexuelle, les ovaires sont gros et contiennent souvent de nombreux ovules vitellogènes à leur surface, en plus des nombreux petits follicules en développement, de tailles variées, pris dans le tissu conjonctif. Chez certaines espèces d'élasmobranches, seul un ovaire est fonctionnel et contribue à la production d'ovules; c'est généralement le cas pour les raies de l'ordre des Myliobatiformes, chez lesquelles l'ovaire gauche est le principal producteur d'ovules matures et l'ovaire droit est rudimentaire. Chez les raies de l'ordre des Rajiformes, les deux ovaires sont généralement fonctionnels.
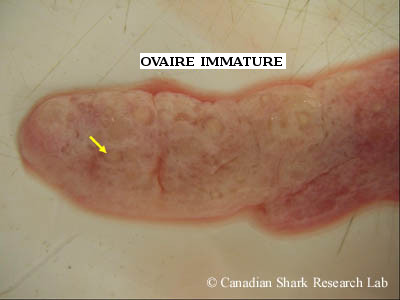
Ovaire d'une raie tachetée (Leucoraja ocellata) immature. Noter le petit ovule blanchâtre et indistinct pris dans le tissu conjonctif.

Ovaire d'une raie tachetée (Leucoraja ocellata) à maturité précoce. Noter les ovules qui commencent à être de différentes tailles et couleurs.
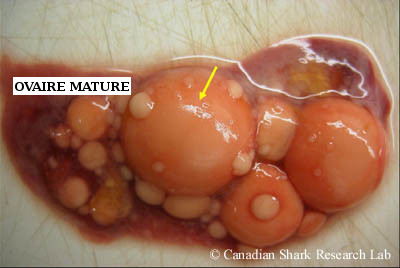
Ovaire d'une raie tachetée (Leucoraja ocellata) mature. Noter les très gros ovules jaune-orange prêts à être fécondés, et les plus petits ovules de différents états vitellogènes et tailles.
Raies (ordre des Myliobatiformes)
Chez les batoïdes dont les cycles de reproduction sont bien définis, comme de nombreuses espèces de raies de l'ordre des Myliobatiformes, le début de la saison de reproduction est marqué par une augmentation du nombre et de la taille des oocytes sur la surface antérieure des ovaires, processus appelé oogénèse. Les oocytes commencent aussi à accumuler progressivement du vitellus (processus appelé synthèse du jaune), passant d'une teinte blanchâtre à une couleur jaune-orange, jusqu'à ce qu'ils soient prêts pour l'ovulation. Ce processus est lent, nécessitant de plusieurs mois à un an chez la plupart des espèces. Au cours de l'ovulation, les plus gros ovules matures sont libérés un par un et passent des ovaires aux ostia (ostium au singulier). Ils vont ensuite dans les oviductes, qui sont de longs tubes parcourant la cavité abdominale et dont la seule fonction est de transporter les ovules. La libération des ovules matures à partir de chaque ovaire stimule la migration des autres ovules matures vers la surface des ovaires, afin qu'il y ait toujours des ovules prêts pour l'ovulation. Après être passé dans l'oviducte antérieur, chaque ovule entre dans la glande nidamentaire (ou coquillière), qui est un renflement de l'extrémité antérieure de l'oviducte. La glande nidamentaire est un organe très spécialisé où le sperme est stocké (chez certaines espèces de batoïdes). C'est là que se produit la fécondation et que chaque ovule fécondé se retrouve ensuite encapsulé dans un fin sac ovigère membraneux, une sorte d'enveloppe, avant la mise bas. La taille de la glande nidamentaire dépend du cycle de reproduction, atteignant son maximum juste avant le début de l'ovulation. Derrière chaque glande nidamentaire se trouve un utérus, un renflement de l'oviducte postérieur. Les deux utérus se réunissent pour former le sinus urogénital, une cavité qui s'ouvre sur le cloaque et grossit au fur et à mesure de la maturation sexuelle.
Chez les raies de l'ordre des Myliobatiformes, l'accouplement, la gestation et la mise bas tendent à se produire de façon synchrone. Le développement des follicules dans les ovaires peut avoir lieu en même temps que la gestation (figure 7), ce qui est caractéristique des élasmobranches femelles qui sont gestantes à longueur d'année, ou encore entre deux gestations, ce qui est le cas de la majorité des raies aux gestations comparativement courtes qui durent plusieurs mois au cours du cycle annuel (figure 8). Les ovaires des femelles inactives sur le plan de la reproduction (c.-à-d. à la fin du cycle de reproduction) ne contiennent que de petits follicules avec peu ou pas de vitellus. La gestation est presque immédiatement suivie de l'accouplement, puis d'une autre gestation. Il y a des exceptions, comme Torpedo marmorata, qui a une gestation d'un an suivie de deux années sans gestation. La mise bas, quant à elle, peut être déclenchée par des facteurs écologiques précis, tels que des changements dans la température ou l'abondance relative des proies pour les nouveau-nés.

Glande nidamentaire droite et utérus antérieur d'une raie mature de l'Atlantique Nord-Ouest. Chez les raies de l'ordre des Rajiformes, les glandes nidamentaires dominent le système reproducteur, tandis que chez les raies de l'ordre des Myliobatiformes, la taille de ces glandes varie visiblement selon la période du cycle de reproduction.
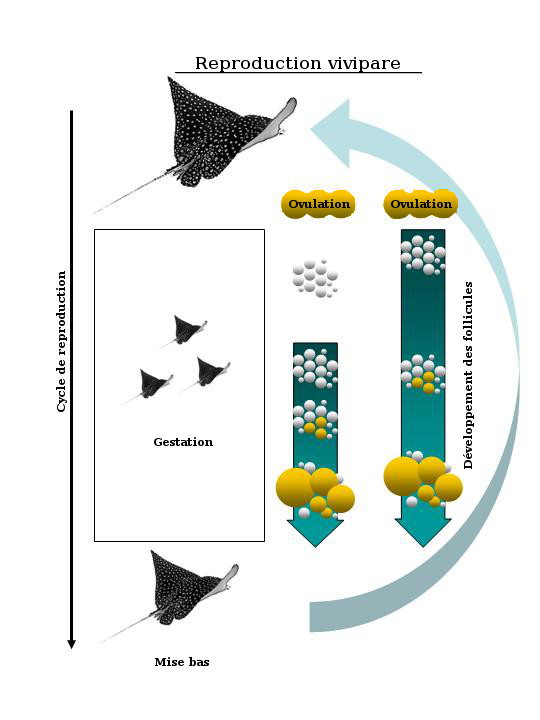
Diagramme illustrant les deux cycles de reproduction vivipares caractéristiques des raies qui ont des gestations à longueur d'année. Dans le premier scénario, le développement des follicules se produit durant la deuxième moitié de la gestation. Dans le second scénario, le développement des follicules se produit pendant la gestation.
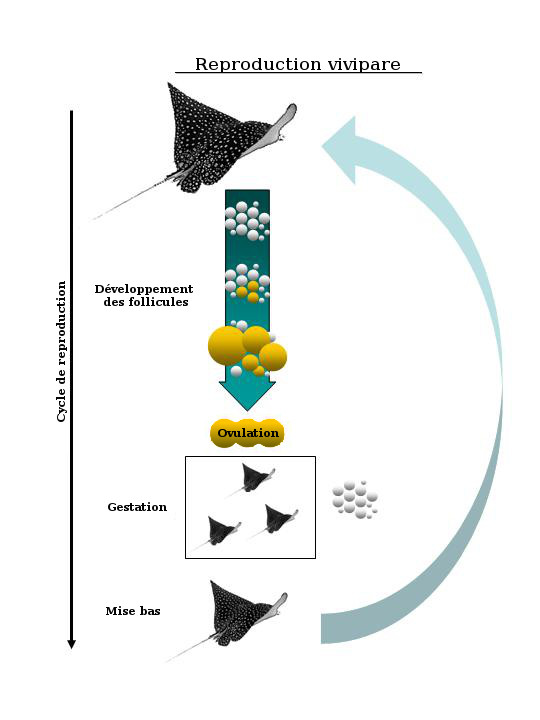
Diagramme illustrant le cycle de reproduction vivipare caractéristique des raies aux gestations plus courtes qui durent plusieurs mois au cours d'un cycle annuel. Le développement des follicules se produit entre les gestations.
Raies (ordre des Rajiformes)
L'anatomie reproductrice interne des raies de l'ordre des Rajiformes est très similaire à celui des raies de l'ordre des Myliobatiformes. Toutefois, contrairement aux raies de l'ordre des Myliobatiformes, les cycles de reproduction saisonniers des espèces d'élasmobranches ovipares (y compris de toutes les espèces de raies de l'ordre des Rajiformes) ne sont d'ordinaire pas clairement marqués par un début et une fin. À l'inverse, la tendance générale est un an de production d'œufs avec des périodes saisonnières ou de pointe pendant lesquelles une plus grande proportion de femelles matures pond leurs œufs. Pour que ces espèces puissent produire des œufs tout au long de l'année, elles présentent en tout temps des paires de follicules dans les ovaires, chaque paire nécessitant au moins plusieurs semaines avant d'atteindre une taille acceptable en vue de l'ovulation. La synthèse du jaune se produit de façon synchrone avec l'oviposition. En d'autres termes, les femelles matures dont le système de reproduction rappelle un « tapis roulant » sont dotées de gros ovules vitellogènes et de nombreux petits ovules non fécondés de diverses tailles dans leurs ovaires tout au long de l'année. De petits ovules sont toujours en développement pour remplacer ceux qui sont encapsulés et pondus (figure 2).
Les raies de l'ordre des Rajiformes produisent des capsules d'œuf, ce qui leur est propre. Chez les deux types de raies, l'encapsulation des ovules se produit dans les glandes nidamentaires. Toutefois, chez toutes les espèces ovipares, la glande nidamentaire domine le système reproducteur, sa taille augmentant lors des périodes de pointe de production d'ovules, puis ne variant que légèrement durant le cycle de reproduction. De plus, en raison de la complexité de la formation des capsules chez les espèces ovipares, la formation débute avant l'ovulation et n'est terminée qu'au tiers ou à la moitié lorsque les ovules fécondés entrent dans la glande nidamentaire. Une fois terminées, les capsules d'œuf produites par les raies sont dotées d'une structure complexe et remarquablement résistante. Elles sont libérées dans l'environnement externe au cours d'un processus appelé oviposition. La formation des capsules d'œuf demeure néanmoins un processus rapide ne durant que de 12 à 24 heures, le durcissement des capsules (ou sclérotisation) se faisant dans l'utérus. Les capsules d'œuf entièrement formées sont alors conservées dans le sinus urogénital pendant plusieurs heures avant l'oviposition; cette étape-là peut durer plusieurs jours.
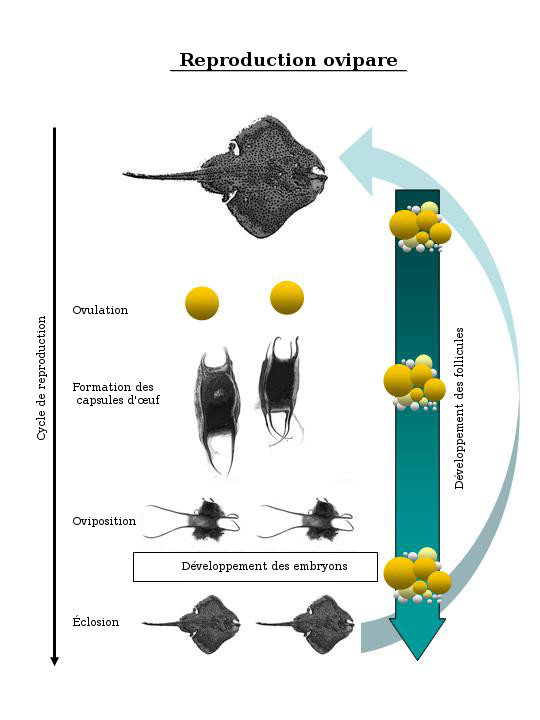
Diagramme illustrant le cycle de reproduction ovipare caractéristique des raies qui présentent un type de reproduction faisant penser à un « tapis roulant ». Le développement des follicules se fait de façon synchrone avec l'oviposition, et une série de paires de follicules de différentes tailles sont présentes en tout temps dans les ovaires.

Système reproducteur d'une raie épineuse (Amblyraja radiata) femelle en pleine période de reproduction. Deux capsules d'œuf entièrement formées sont visibles dans les utérus derrière les glandes nidamentaires. Le sinus urogénital est bien dilaté.

Système reproducteur d'une raie épineuse (Amblyraja radiata) femelle en pleine période de reproduction. Le système reproducteur de droite a été partiellement retiré de la cavité abdominale pour montrer la glande nidamentaire de droite et la capsule d'œuf entièrement formée présente dans l'utérus.
Pour obtenir un aperçu plus détaillé des œufs après l'oviposition, veuillez consulter la section Capsules d'œuf du site Web. Pour en apprendre davantage sur la façon dont la maturité et les cycles de reproduction sont étudiés chez les raies, veuillez consulter la section Recherche sur les raies du site Web.
- Date de modification :